Obesity, diabetes and metabolic syndrome are major public health problems and significant risk factors for heart disease in the United States and world-wide. Sah lab is currently focused on studying ion channels as regulators of cellular metabolism in numerous tissues and cell types, including adipose tissue, pancreatic islet cells, skeletal muscle, liver and myocardium. Specifically, our research interests focus on intracellular signaling of transient receptor potential channels (including TRPV3, TRPV4 and TRPM7), the recently identified essential component of the volume regulatory anion channel SWELL1 (LRRC8a), and mechanosensitive ion channel (Piezo1) as they relate to obesity and metabolism.
To do this we combine cellular electrophysiology, calcium imaging (GCaMP6) and novel genetic techniques (including transient and stable lenti/AAV-shRNA-mediated knockdown and CRISPR/cas9-mediated knockout) in cultured cells (mouse and human) and freshly isolated primary cells (adipocytes, islets, endothelium, skeletal muscle fibers, cardiomyocytes). Genetic loss-of-function mouse models and knock-in approaches are also used to examine the contributions of these ion channels in vivo and in disease settings.

SWELL1-LRRC8 signaling in adipocytes: Can fat “feel” fat?
Obesity is a major public health problem, predisposing to high cholesterol, diabetes and hypertension and inflicting healthcare costs over 100 billion dollars in the U.S. alone. It is characterized by a tremendous increase in adipose tissue that is in large part due to massive volumetric expansion of the constituent adipocytes1, 2. This adipocyte expansion has been long associated with metabolic disease and insulin resistance1, 3-6, implicating adipocyte-size dependent regulation of insulin signaling and growth.
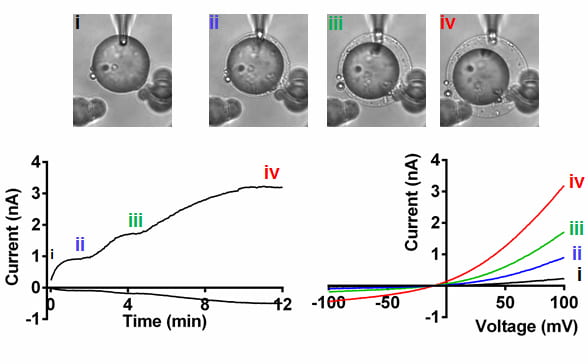
Adipocyte patch-clamp reveals a swell-activated current.
We recently discovered that SWELL1 (LRRC8a), a member of the Leucine Rich Repeat Containing protein family, is a required component of a volume-sensitive and putatively mechanosensitive ion channel/signaling molecule that is activated in the setting of adipocyte hypertrophy and regulates adipocyte size, insulin signaling and systemic glycaemia via a novel SWELL1-PI3K-AKT2-GLUT4 signaling axis
Adipocyte-specific SWELL1 ablation disrupts insulin-PI3K-AKT2 signaling, inducing insulin resistance and glucose intolerance in vivo7. These data identify SWELL1 as a component of a cell-autonomous sensor of adipocyte size and a positive regulator of adipocyte insulin signaling, growth and glucose homeostasis, particularly in the setting of obesity. Thus, SWELL1 may be a component of the molecular mechanosensor responsible for the mechanosensitive nature of adipocytes described by others previously8-11.
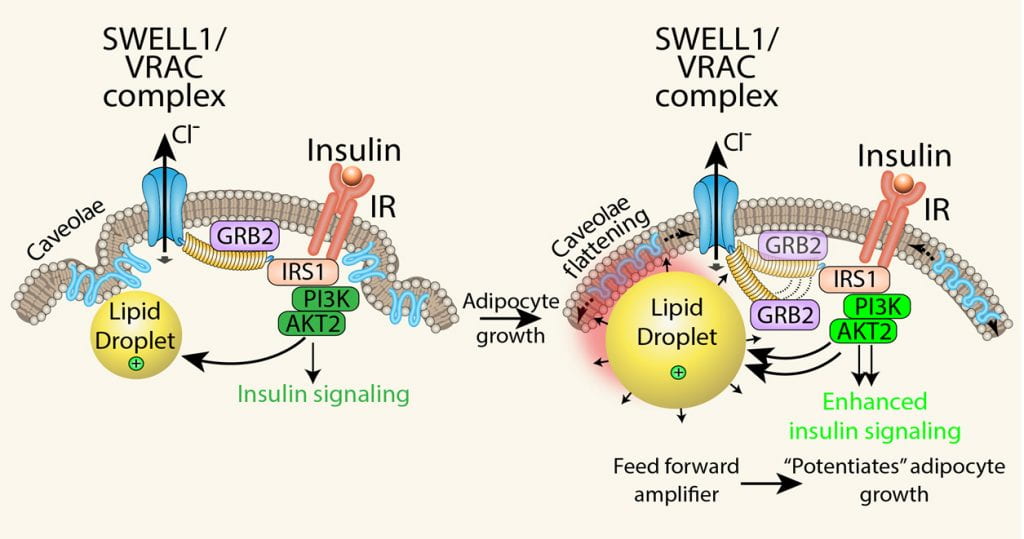
Adipocyte SWELL1/VRAC complex regulates insulin-PI3K-AKT2 signaling and senses mechanical forces via caveolae flattening to tune insulin- AKT2 signaling by modulating GRB2-IRS1 interactions. Working model: adipocyte expansion activates the SWELL1/VRAC complex to disinhibit insulin-PI3K-AKT2 via GRB2, and thereby support lipogenesis and continued adipocyte growth, in a feed-forward manner, potentiating adipocyte hypertrophy.
SWELL1-LRRC8a mediated glucose-sensing in pancreatic β-cells
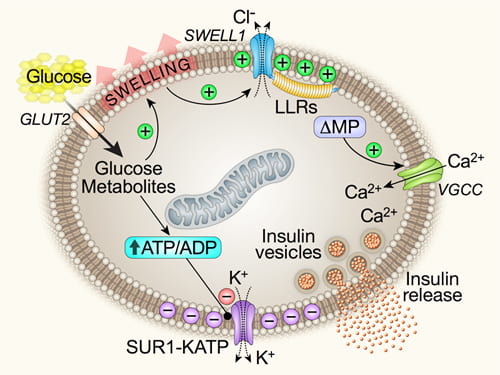
Working model: SWELL1 is a virtual glucose sensor regulating β-cell excitability and systemic glycaemia. GLUT2, glucose transporter; ΔMP, membrane depolarization; VGCC, voltage-gated calcium channel.
Insulin secretion from the pancreatic b-cell is triggered by Ca2+ influx through voltage-gated Ca2+ channels (VGCC) to induce insulin vesicle fusion with the β-cell plasma membrane. Activation of VGCC depends on the b-cell membrane potential, which is in turn mediated by the balance of depolarizing (excitatory) and hyperpolarizing (inhibitory) ionic currents12. While much attention has focused on inhibitory hyperpolarizing currents there is little knowledge about the requisite excitatory currents responsible for depolarizing the β-cell, including the molecular identity of these currents12, 13. One candidate for a depolarizing current is a chloride conductance (Cl–) known as the volume-regulatory anion current (VRAC) or ICl,SWELL12, 14-30. However, the molecular identity of the channel responsible for this current in β-cells had been elusive for decades.
We recently reported that SWELL1 (LRRC8a)31-34, a member of the Leucine Rich Repeat Containing protein family, is required for ICl,SWELL in β-cells35. SWELL1-mediated ICl,SWELL activates upon β-cell swelling induced by glucose import29, and this generates a depolarizing current contributing to VGCC activation, thereby regulating insulin secretion and systemic glycemia35. In this model, the hyperpolarizing K+ conductances act as “brakes” on β-cell excitability and insulin secretion, while SWELL1-mediated ICl,SWELL is the “accelerator”—promoting β-cell excitability in response to glucose-mediated β-cell swelling. We propose that it is the balance between ICl,SWELL and IKATP that is critical in regulating β- cell membrane potential.
References
1. Farnier, C., et al. Adipocyte functions are modulated by cell size change: potential involvement of an integrin/ERK signalling pathway. Int J Obes Relat Metab Disord 27, 1178-1186 (2003).
2. Kim, S.M., et al. Loss of white adipose hyperplastic potential is associated with enhanced susceptibility to insulin resistance. Cell metabolism 20, 1049-1058 (2014).
3. Salans, L.B., Knittle, J.L. & Hirsch, J. The role of adipose cell size and adipose tissue insulin sensitivity in the carbohydrate intolerance of human obesity. J Clin Invest 47, 153-165 (1968).
4. Heinonen, S., et al. Adipocyte morphology and implications for metabolic derangements in acquired obesity. Int J Obes (Lond) (2014).
5. Weyer, C., Foley, J.E., Bogardus, C., Tataranni, P.A. & Pratley, R.E. Enlarged subcutaneous abdominal adipocyte size, but not obesity itself, predicts type II diabetes independent of insulin resistance. Diabetologia 43, 1498-1506 (2000).
6. Lonn, M., Mehlig, K., Bengtsson, C. & Lissner, L. Adipocyte size predicts incidence of type 2 diabetes in women. FASEB J 24, 326-331 (2010).
7. Zhang, Y., et al. SWELL1 is a regulator of adipocyte size, insulin signalling and glucose homeostasis. Nature cell biology 19, 504-517 (2017).
8. Ben-Or Frank, M., Shoham, N., Benayahu, D. & Gefen, A. Effects of accumulation of lipid droplets on load transfer between and within adipocytes. Biomech Model Mechanobiol (2014).
9. Shoham, N., et al. Adipocyte stiffness increases with accumulation of lipid droplets. Biophys J 106, 1421-1431 (2014).
10. Shoham, N., et al. Static mechanical stretching accelerates lipid production in 3T3-L1 adipocytes by activating the MEK signaling pathway. Am J Physiol Cell Physiol 302, C429-441 (2012).
11. Pellegrinelli, V., et al. Human adipocyte function is impacted by mechanical cues. The Journal of pathology 233, 183-195 (2014).
12. Yang, S.N., et al. Ionic mechanisms in pancreatic beta cell signaling. Cell Mol Life Sci 71, 4149-4177 (2014).
13. Rorsman, P., Eliasson, L., Kanno, T., Zhang, Q. & Gopel, S. Electrophysiology of pancreatic beta-cells in intact mouse islets of Langerhans. Prog Biophys Mol Biol 107, 224-235 (2011).
14. Best, L., Miley, H.E. & Yates, A.P. Activation of an anion conductance and beta-cell depolarization during hypotonically induced insulin release. Exp Physiol 81, 927-933 (1996).
15. Kinard, T.A. & Satin, L.S. An ATP-sensitive Cl- channel current that is activated by cell swelling, cAMP, and glyburide in insulin-secreting cells. Diabetes 44, 1461-1466 (1995).
16. Beauwens, R., et al. Stimulus-secretion coupling of hypotonicity-induced insulin release in BRIN-BD11 cells. Endocrine 30, 353-363 (2006).
17. Best, L. Glucose-sensitive conductances in rat pancreatic beta-cells: contribution to electrical activity. Biochimica et biophysica acta 1468, 311-319 (2000).
18. Best, L. Evidence that glucose-induced electrical activity in rat pancreatic beta-cells does not require KATP channel inhibition. J Membr Biol 185, 193-200 (2002).
19. Best, L. How does glucose evoke electrical activity in the pancreatic beta-cell? Diabetes Metab 28, 3S18-24; discussion 13S108-112 (2002).
20. Best, L. Study of a glucose-activated anion-selective channel in rat pancreatic beta-cells. Pflugers Arch 445, 97-104 (2002).
21. Best, L. Glucose-induced electrical activity in rat pancreatic beta-cells: dependence on intracellular chloride concentration. J Physiol 568, 137-144 (2005).
22. Best, L., Brown, P.D., Sener, A. & Malaisse, W.J. Electrical activity in pancreatic islet cells: The VRAC hypothesis. Islets 2, 59-64 (2010).
23. Best, L., Brown, P.D., Sheader, E.A. & Yates, A.P. Selective inhibition of glucose-stimulated beta-cell activity by an anion channel inhibitor. J Membr Biol 177, 169-175 (2000).
24. Best, L., Miley, H.E., Brown, P.D. & Cook, L.J. Methylglyoxal causes swelling and activation of a volume-sensitive anion conductance in rat pancreatic beta-cells. J Membr Biol 167, 65-71 (1999).
25. Best, L., Sheader, E.A. & Brown, P.D. A volume-activated anion conductance in insulin-secreting cells. Pflugers Arch 431, 363-370 (1996).
26. Best, L. & Yates, A.P. Electrophysiological effects of osmotic cell shrinkage in rat pancreatic beta-cells. Islets 2, 303-307 (2010).
27. Best, L., Yates, A.P., Decher, N., Steinmeyer, K. & Nilius, B. Inhibition of glucose-induced electrical activity in rat pancreatic beta-cells by DCPIB, a selective inhibitor of volume-sensitive anion currents. Eur J Pharmacol 489, 13-19 (2004).
28. McGlasson, L., Best, L. & Brown, P.D. The glucokinase activator GKA50 causes an increase in cell volume and activation of volume-regulated anion channels in rat pancreatic beta-cells. Mol Cell Endocrinol 342, 48-53 (2011).
29. Miley, H.E., Sheader, E.A., Brown, P.D. & Best, L. Glucose-induced swelling in rat pancreatic beta-cells. J Physiol 504 ( Pt 1), 191-198 (1997).
30. Sheader, E.A., Brown, P.D. & Best, L. Swelling-induced changes in cytosolic [Ca2++] in insulin-secreting cells: a role in regulatory volume decrease? Mol Cell Endocrinol 181, 179-187 (2001).
31. Qiu, Z., et al. SWELL1, a Plasma Membrane Protein, Is an Essential Component of Volume-Regulated Anion Channel. Cell 157, 447-458 (2014).
32. Voss, F.K., et al. Identification of LRRC8 heteromers as an essential component of the volume-regulated anion channel VRAC. Science 344, 634-638 (2014).
33. Syeda, R., et al. LRRC8 Proteins Form Volume-Regulated Anion Channels that Sense Ionic Strength. Cell 164, 499-511 (2016).
34. Deneka, D., Sawicka, M., Lam, A.K.M., Paulino, C. & Dutzler, R. Structure of a volume-regulated anion channel of the LRRC8 family. Nature (2018).
35. Kang, C., et al. SWELL1 is a glucose sensor regulating beta-cell excitability and systemic glycaemia. Nat Commun 9, 367 (2018).